Louis Bernatchez ,Julian Dodson , Helga Guderley (U. Laval)
L’adaptation locale est un processus qui augmente la fréquence des traits adaptatifs dans une population. Un trait adaptatif est un attribut qui augmente la survie ou le succès reproducteur des individus qui l’expriment. L’objectif de ce projet est d’élucider les bases génétiques de traits morphologiques et physiologiques reliés au fitness chez le saumon atlantique. Cette recherche représente le premier effort visant à tester d’une manière étayée et rigoureuse les bases génétiques du concept de l’adaptation locale chez le saumon atlantique, ce qui contribuera à améliorer les mesures de conservation chez cette espèce et à mieux comprendre et prédire les changements génétiques induits par certaines manipulations de populations faites par l’homme, notamment, l’établissement de nouvelles populations par introduction dans de nouveaux habitats ou encore l’échappement de poissons d’élevage et l’hybridation avec les populations naturelles.
Objectifs
spécifiques |
(1)Variation latitudinale de la diversité allélique d’un gène d’immunocompétence : co-évolution hôte-pathogène et adaptation locale chez le saumon atlantique?
(2) Comparaison de profils d’expression de gènes entre une lignée domestique de saumon et sa population fondatrice.
(3) Comparaison des profils d’expression des gènes (a) entre la progéniture de saumons d’une population nouvellement formée sur la rivière Sainte-Marguerite (5 générations) et celle de saumons de sa population fondatrice; (b) entre la progéniture de mâles anadromes ou à maturité sexuelle précoce chez ces 2 populations.
(4) Comparaison des traits morphologiques et métaboliques et de profils d'expression reliés au déterminisme de la croissance, chez ces 2 populations de saumon atlantique présentant des taux différents de maturité sexuelle précoce et quantification de l'héritabilité de ces traits ;
(5) Préférence d'habitat des tacons de saumon atlantiqueà maturité sexuelle précoce.
Quelques résultats
(1) Variation
latitudinale de la diversité allélique d’un gène d’immunocompétence
: co-évolution hôte-pathogène et adaptation
locale chez le saumon atlantique? |
|||
|
|||
|
|||
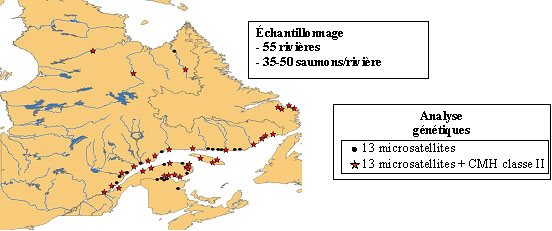
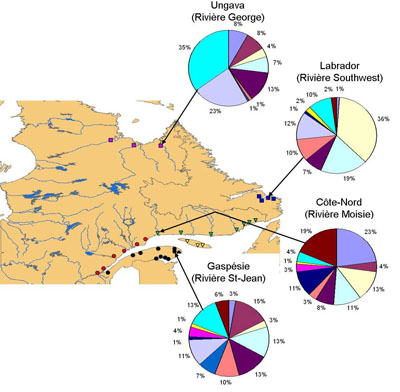
| Les populations de saumon atlantique étudiées
possèdent entre 9 et 14 allèles différents
au gène CMH II, ce qui représente un polymorphisme
relativement élevé. La diversité génétique des populations de saumons au gène CMH est élevée dans la plupart des rivières de la Gaspésie, alors que le Labrador et l’Ungava sont des régions où la diversité génétique est plus faible à ce gène. |
|||
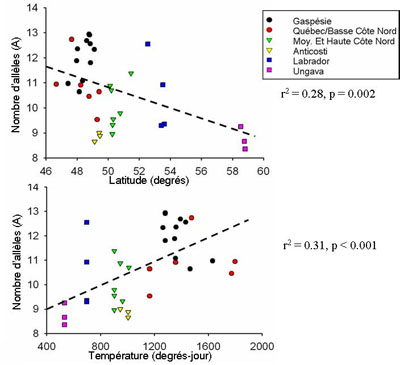
Les résultats obtenus indiquent une
diminution de la diversité génétique au gène
CMH classe II avec la latitude (r2 = 0.28, p = 0.002) et avec la
diminution de la température (r2 = 0.31, p < 0.001). |
|||
|
|||
(2) Comparaison de profils d’expression de gènes entre une lignée domestique de saumon et sa population fondatrice |
||
|
||
Cinq à sept
générations de sélection artificielle visant,
entre autres choses, une augmentation du taux de croissance et
une diminution de la proportion de madeleineaux chez le saumon
atlantique, ont suffit pour entraîner des modifications
moyennes de 20% du niveau d'expression d’au moins 1.7 et
1.4% des gènes exprimés dans les souches d'élevage
norvégienne et canadienne respectivement, et ce, pour
différents groupes de fonctions. Certains gènes
montraient en outre des changements parallèles dans les
souches des deux pays. Ces résultats supportent donc l’hypothèse
voulant que l’hybridation éventuelle entre saumons
d’élevage et sauvages puisse altérer négativement
le pool génique des populations naturelles et en diminuer
le fitness. |
 Alevins
de saumons peu après l'éclosion avec, en arrière
plan, l'image d'une bio-puce Photo des alevins © Normand Bergeron (extraite du documentaire "Jusqu'à la mer"); Image de bio-puce et montage © Christian Roberge |
|
|
||
(3) Comparaison des profils d’expression des gènes (a) entre la progéniture de saumons d’une population nouvellement formée sur la rivière Sainte-Marguerite (5 générations) et celle de saumons de sa population fondatrice; (b) entre la progéniture de mâles anadromes ou à maturité sexuelle précoce chez ces 2 populations |
|||
|
L’installation, en 1981,
d’une passe migratoire à l’emplacement d’une
chute sur la branche Nord-Est de la rivière Sainte-Marguerite
a rendu accessible un segment de rivière jadis inoccupé par
l’espèce. Depuis, après 5 générations,
une sous-population génétiquement distincte, et
en voie d'isolement reproducteur, s’y est établie. De plus, la proportion de mâles à maturité sexuelle précoce est plus importante en amont de la chute qu’en aval. |
 |
|
|
|||
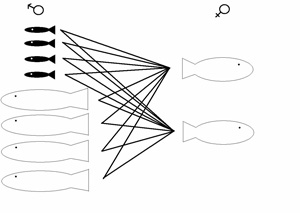 |
2
séries de croisements ont été effectuées
pour 2 années consécutives (pisciculture de Tadoussac)
avec des géniteurs (mâles anadromes ou précoces
et femelles anadromes) capturés en amont (série 1)
et en aval (série
2) de la passe migratoire. Ces croisements permettent de quantifier par analyse de variance les effets de populations, maternels et de tactiques de reproduction sur les profils d’expression. Les profils d’expression sont analysés à l'aide de bio-puces aux stades de vie embryonnaire et juvénile. |
||
|
|||
(4) Comparaison
des traits morphologiques et métaboliques et de
profils d'expression reliés
au déterminisme de la croissance chez
ces 2 populations de saumon atlantique présentant
des taux différents de maturité sexuelle précoce
et quantification de l'héritabilité de ces
traits |
Résultats à venir
|
(5) Préférence
d'habitat des tacons de saumon atlantiqueà maturité sexuelle
précoce |
Résultats à venir |